


To describe the antimicrobial activity of β-defensin-2 produced in the mammary gland and secreted in human breast milk.
MethodsThe peptide production was performed by DNA cloning. β-defensin-2 levels were quantified in 61 colostrum samples and 39 mature milk samples from healthy donors, by an indirect enzyme-linked immunosorbent assay (ELISA). Using halo inhibition assay, this study assessed activity against seven clinical isolates from diarrheal feces of children between 0 and 2 years of age. The activity of β-defensin-2 against three opportunistic pathogens that can cause nosocomial infections was determined by microdilution test.
ResultsThe peptide levels were higher in colostrum (n=61) than in mature milk samples (n=39), as follows: median and range, 8.52 (2.6-16.3) μg/ml versus 0.97 (0.22-3.78), p<0.0001; Mann-Whitney test. The recombinant peptide obtained showed high antimicrobial activity against a broad range of pathogenic bacteria. Its antibacterial activity was demonstrated in a disk containing between 1–4μg, which produced inhibition zones ranging from 18 to 30mm against three isolates of Salmonella spp. and four of E. coli. β-defensin-2 showed minimum inhibitory concentrations (MICs) of 0.25μg/mL and 0.5μg/mL for S. marcescen and P. aeruginosa, respectively, while a higher MIC (4μg/mL) was obtained against an isolated of multidrug-resistant strain of A. baumannii.
ConclusionsTo the authors’ knowledge, this study is the first to report β-defensin-2 levels in Latin American women. The production and the activity of β-defensin-2 in breast milk prove its importance as a defense molecule for intestinal health in pediatric patients.
Descrever a atividade antimicrobiana da defensina-beta 2 na glândula mamária e secretada no leite materno humano.
MétodosA produção de peptídeos foi realizada por clonagem de DNA. Os níveis de defensina-beta 2 foram quantificados em 61 amostras de colostro e 39 de leite maduro de doadoras saudáveis pelo teste ELISA indireto. Por um ensaio de halo de inibição, avaliamos a atividade contra sete isolados clínicos diarreicos de crianças entre 0 e 2 anos. A atividade da defensina 2 contra três patógenos oportunistas que podem causar infecções nosocomiais foi determinada pelo teste de microdiluição.
ResultadosOs níveis de peptídeos estavam significativamente maiores nas amostras de colostro (n=61) que de leite maduro (n=39), como segue: 8,52 (2,6-16,3μg/mL) mediana e faixa em comparação a 0,97 (0,22-3,78), p<0,0001; teste de Mann-Whitney. O peptídeo recombinante foi obtido da alta atividade antimicrobiana demonstrada contra uma ampla gama de bactérias patogênicas. Sua atividade antibacteriana foi demonstrada em um disco contendo entre 1-4μg, que produziu zonas de inibição entre 18 e 30mm contra três isolados de Salmonella spp. e quatro de E. coli. A defensina-beta 2 demonstrou concentrações inibitórias mínimas (CIMs) de 0,25μg/mL e 0,5μg/mL para S. marcescen and P. aeruginosa, ao passo que uma CIM maior (4μg/mL) foi obtida contra um isolado de cepa multirresistente de A. baumannii.
ConclusõesAté onde sabemos, este estudo é o primeiro a relatar níveis de defensina em mulheres da América Latina. A produção e a atividade da defensina 2 no leite materno comprovam sua importância como uma molécula de defesa para a saúde intestinal em pacientes pediátricos.
The World Health Organization reports that 6.6 million children under the age of 5 died in 2012. Moreover, 44% of those deaths occurred during the neonatal period. From the end of the neonatal period through the first 5 years of life, the main causes of death are pneumonia, diarrhea, and malaria. Malnutrition is the underlying contributing factor in approximately 45% of all child deaths, making children more vulnerable to severe diseases.1 Over half of these early child deaths are due to conditions that could be prevented or treated with access to simple, affordable interventions; one of them is breastfeeding.
Solely during lactancy, humans confront the challenge of survival by means of breast milk. Human milk is associated with protection against diarrheal diseases, respiratory-tract infections, and necrotizing enterocolitis (NEC).2–5 Protection against infection in breastfed infants appears to occur through a variety of complementary acquired and innate defense factors found in human milk, including oligosaccharides and their glycoconjugates,6 and antimicrobial peptides (AMPs).7,8
Among AMPs, the defensins family constitutes an important part of the innate immune response in fluids secretion. Increasing evidence suggests the importance of defensins in the immune response. Results in recent studies on the homeostatic and disease-fighting activities of human defensins point to their key relevance in several pediatric disorders. For example, high β-defensin-2 (hBD2) concentrations reflecting strong intestinal immune responses were associated with moderate courses of NEC.9 However, infants with severe NEC showed no increase in fecal hBD2 concentrations before and during the disease. These studies suggest that a specific deficiency of innate defense activation in extremely-low birth weight (ELBW) infants rather than an impaired intestinal epithelial barrier leads to a more severe course of NEC.9 In this context, the intake of hBD2 from breast milk gains importance. Notably, intestinal hBD2 levels correspond to both defensin in breast milk7,8,10 and secretions of intestinal epithelial cells. The latter are induced by catalytic antibodies contained in breast milk, acting through protease-activated receptor-2 present in the plasma membrane.11
The concentrations of total and specific components of human breast milk vary between mothers, diurnally, based on infant's gestational age, and over the course of lactation.10,12 Presence of intact hBD2 has also been reported in infant feces, which shows that hBD2 tends to withstand the digestive processes of the gastrointestinal tract.13 Indeed, it has been proposed that defensin quantification in feces could be an effective biomarker of intestinal health and disease in pediatric population.14 The resistance of hBD2 to digestion would increase its availability, which would allow to protect the entire mucosal surface of the gastrointestinal tract.
This study had a two-fold aim. First, to quantify the hBD2 in breast milk from Venezuelan donors. Second, to characterize the antimicrobial activity of BD2 against both bacterium associated with enterocolitis in infants and against Gram-negative bacteria associated with nosocomial infections.
Materials and methodsMaterialsThe TOPO® TA Cloning® kit for sequencing and the pET directional TOPO® expression kit were purchased from Invitrogen (Invitrogen Corp., Carlsbad, Califórnia, USA). All other reagents were of analytical reagent grade.
Human milk collection and preparationAn informed consent from all donors involved in this study was obtained prior to obtaining and processing the samples. None of the donors in the study had history of rheumatologic, respiratory, cardiovascular, or gastrointestinal diseases. The inclusion criteria were healthy lactating mothers with term delivery. The exclusion criteria were mothers who had had cesarean deliveries, were receiving antibiotic treatment on suspicion of infection, or with a history of autoimmune diseases. Samples from day one to day seven after childbirth were denoted as colostrum, whereas samples collected beyond the tenth day of birth were termed mature milk.
This study obtained 61 colostrum and 39 mature samples. All samples were collected using a standardized protocol approved by the Bioethics Committee of the Clinical University Hospital at Universidad Central de Venezuela (UCV). Samples were collected from healthy human mothers (18 to 30 years old) with support from the milk bank service of the Clinical University Hospital. Prior to extraction, the nipples were cleaned and swabbed with warm sterile water.
Samples were collected from one breast, and centrifuged at 4°C (1,000×g for 15min) so that samples had a pellet of cells at the bottom and a fat layer at the top. Whey supernatant in the middle was aliquoted and frozen at −80°C. When ready for use, milk was thawed at 4°C.
Cells culturesCultured SKBR3 cells (human breast adenocarcinoma cell line, HTB-30 cat., No. 11965-092 American Type Culture Collection (ATCC), Manassas, USA).
Bacterial strainsClinical isolates: Acinetobacter baumannii (multidrug-resistant strain isolated from a patient suffering nosocomial infection of Caracas University Hospital-UCV) was kindly provided by Dr. G. Alonso (Instituto de Biología Experimental-UCV). In total, seven enteropathogens from children's diarrheal samples were tested, three belonging to genus Salmonella (Salmonella paratyphi, Salmonella arizona, Salmonella sp.) and four different isolates of E. coli. All were identified with standard bacteriological assays using a Vitek® 32 and GNI card (BioMérieux Inc., Missouri, USA).
The Pseudomonas aeruginosa ATCC 276532 strain was courtesy of Dr. Takiff of Molecular Genetics Laboratory- IVIC. Escherichia coli ATCC 25922 was obtained from ATCC.
AnimalsFive-month-old brown hens were bought from a local farmer.
Reverse transcription polymerase chain reaction (RT-PCR)SKBR3 cells were grown to 80% confluence. Cells were then treated for 4h with lipopolysaccharide (LPS) (1μg/mL), as described previously.11 RNA was extracted from SKBR3 cells using Trizol® (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. BD2 cDNA was amplified by RT-PCR using sense and antisense hBD2 primers, as previously reported.11 Directional cloning of PCR product was achieved using designed primers with particular restriction sites.
hBD2S:CCCGGGCATGAGGGTCTTGTATCTCCTCTTC
hBD2R:GAATTCGGAGCCCTTTCTGAATCCGCA; (255bp)
The resulting PCR products were purified and encoding full-length gene hBD2 from SKBR3 cells was cloned in pET 100 vector (Champion pET directional TOPO expression and cloning kit, Invitrogen). The fusion proteins were expressed in an E. coli (Strain BL21 DE3) system according to manufacturer's instructions. The recombinant fusion-protein was used in both antimicrobial activity assays. It also was used to produce antibodies from hens’ egg yolks.
Production and purification of antibodies from egg yolks for recombinant hBD2The antigen 100μg (recombinant hBD2) was diluted in PBS (pH 7.2) and emulsified with an equal volume of incomplete Freund's adjuvant. Suspension was injected subcutaneous in the birds. Further injections of the protein, were given to the hens 7, 21, 31, and 42 days later. IgY was purified from individual eggs using a commercial Chicken IgY Purification kit (Pierce Biotechnology, Cat N° 44918, Illinois, USA) following manufacturer instructions. Prior to the immunization, IgY was purified in order to obtain pre-immune IgY to be used as negative control in the enzyme-linked immunosorbent assay (ELISA) standardization.
hBD2 quantification by indirect ELISAAn indirect ELISA was designed for quantification of β-defensin 2. The 96-well immunoplates (MaxisorpTM; Nunc) were coated overnight with clarified milk samples and with the β-defensin 1 or 2 peptides, used as standards. Commercial recombinant Human BD-1 and BD-2 (Cat N° 300-51A and 300-49; Peprotech México) were used. Standards and samples were diluted into carbonate coating buffer, and plates were blocked during 4h in 200mL blocking buffer. Polyclonal IgY chicken anti-human β-defensin-2 was diluted in blocking buffer 1:1000 and added (100μL). Two hours later, secondary rabbit anti-immunoglobulin Y (IgY) peroxidase-conjugate (1:5000 dilution) in blocking buffer was added (100μL) and incubated for 1.5h. The colorimetric reaction was performed as described previously.11 As a control, samples in each plate were run with pre-immune IgY or without primary antibodies to ascertain any unspecific binding. The defensin concentration of samples was computed by interpolating three standard curves constructed via serial twofold dilutions of the recombinant hBD2. The optimal sample dilution was established by titration and varied between donors. The control tests demonstrated that the polyclonal used for β-defensins-2 were selective. Thus, the IgY anti β-defensin 2 did not react with β-defensin 1.
Antimicrobial activity assaysThe assayed strains were grown in liquid Luria-Bertani (LB) medium to OD600 ∼1.0. The resulting suspensions were diluted to a concentration of ∼104 CFU per mL. A 100μL aliquot of LB was dispensed into 96-well plates.15 The antibacterial activity of hBD2 was evaluated with serially diluted peptide to obtain concentrations of 8; 4; 2; 1; 0.5; 0.25 and 0.125μg/mL. A growth control containing no hBD2 and a sterility control without inoculum were also included for each strain. Plates were incubated at 35°C and measurements of turbidity at 600nm using the Synergy HT multidetection microplate reader (Synergy HT, Bio-Tek Instruments, Vermont, USA). In a single experiment, each hBD2 concentration was tested in triplicate. Each microorganism was analyzed in at least three independent experiments. Statistical analysis and dose response curves were obtained using the K4 software provided by Synergy HT (Bio-Tek Instruments, Vermont, USA). Minimum inhibitory concentrations (MICs) were determined from the growth inhibition profiles curves, measured with an absorbance of OD6 00nm, and represent the lowest concentration of the hBD2 that inhibited the bacterial growth by 90% at 24h of culture.
Antimicrobial susceptibility by disk diffusion testsIn vitro susceptibility of clinical isolates to defensin could be roughly determined by disk diffusion tests.16 Kanamycin was used as an internal control. Disks were prepared using a stock solution of antibiotic adding antimicrobial solution to each paper disk, using Filter paper sheets from BioRad (Thick blot paper catalog No. 170-3956). They were prepared within one hour prior to the application to the plates, allowing approximately 30minutes for antibiotic absorption. Prepared disks were not stored and were dry before application.
After 24h of incubation, the inhibition zones were measured. Diameter of the areas of inhibition was converted to the categories of susceptible, intermediate, or resistant (S, I, or R, respectively) according to standard tables. The interpretation, breakpoints, and categorization scheme followed the Clinical and Laboratory Standard Institute (CLSI) recommendations.17
Statistical analysisGraphical presentation and statistical analysis of the data were performed using GraphPad Prism (version 4.0) software (GraphPad Software Inc., California, USA). The Mann-Whitney U test was used for comparison between groups. Statistical significance was set to p<0.05.
ResultsExpression of the recombinant hBD2 peptideThe recombinant hBD2 peptide was produced from mammary epithelial cell cDNA based on the known nucleotide sequence of hBD2 (also known as DEFB4A). Its identity was set by DNA sequence analysis. This recombinant peptide was used to produce polyclonal IgY, whose antibodies were used in indirect ELISA for detection and quantification of human β-defensin in human milk samples.
Quantification of hBD2 in human breast milk samplesA total of 100 healthy Venezuelan mothers were included in this study. The peptide levels were significantly higher in colostrum (8.52 [2.6-16.3μg/mL]) than in mature milk samples (0.97 [0.22-3.78]; p<0.0001; Mann-Whitney test). The hBD2 quantification in human milk samples showed inter-individual variability as well as in each stage of lactation (Fig. 1).
 median and range, than in mature milk samples 0.97 (0.22-3.78). A Mann-Whitney U test was used for comparison between groups, P<0.0001.")
Defensin levels found in the breast milk samples from the 100 donors. The horizontal lines indicate the median of defensin concentration in colostrum and mature samples. The peptide levels were significantly higher in colostrum: 8.52 (2.6-16.3μg/mL) median and range, than in mature milk samples 0.97 (0.22-3.78). A Mann-Whitney U test was used for comparison between groups, P<0.0001.
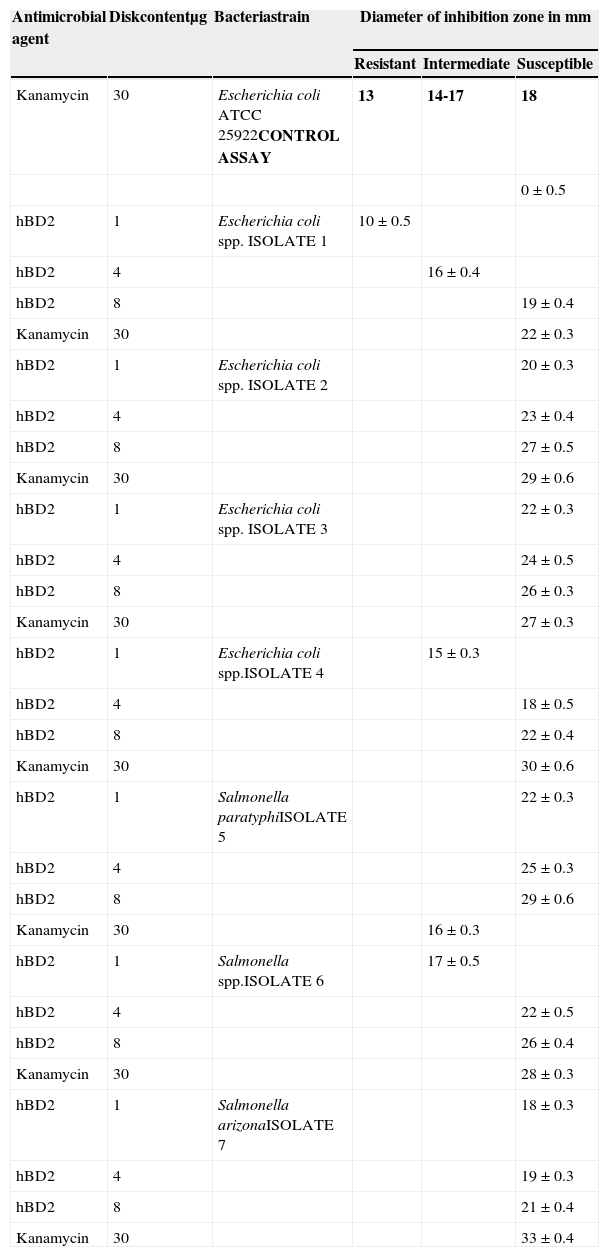
The recombinant peptide was analyzed for anti-enteropathogens activity. Results of these experiments are shown in Table 1 and Fig. 2.
Minimum inhibitory concentrations of human beta defensin-2 peptide against seven clinical isolate strains of Salmonella and E.coli.
| Antimicrobial agent | Diskcontentμg | Bacteriastrain | Diameter of inhibition zone in mm | ||
|---|---|---|---|---|---|
| Resistant | Intermediate | Susceptible | |||
| Kanamycin | 30 | Escherichia coli ATCC 25922CONTROL ASSAY | 13 | 14-17 | 18 |
| 0±0.5 | |||||
| hBD2 | 1 | Escherichia coli spp. ISOLATE 1 | 10±0.5 | ||
| hBD2 | 4 | 16±0.4 | |||
| hBD2 | 8 | 19±0.4 | |||
| Kanamycin | 30 | 22±0.3 | |||
| hBD2 | 1 | Escherichia coli spp. ISOLATE 2 | 20±0.3 | ||
| hBD2 | 4 | 23±0.4 | |||
| hBD2 | 8 | 27±0.5 | |||
| Kanamycin | 30 | 29±0.6 | |||
| hBD2 | 1 | Escherichia coli spp. ISOLATE 3 | 22±0.3 | ||
| hBD2 | 4 | 24±0.5 | |||
| hBD2 | 8 | 26±0.3 | |||
| Kanamycin | 30 | 27±0.3 | |||
| hBD2 | 1 | Escherichia coli spp.ISOLATE 4 | 15±0.3 | ||
| hBD2 | 4 | 18±0.5 | |||
| hBD2 | 8 | 22±0.4 | |||
| Kanamycin | 30 | 30±0.6 | |||
| hBD2 | 1 | Salmonella paratyphiISOLATE 5 | 22±0.3 | ||
| hBD2 | 4 | 25±0.3 | |||
| hBD2 | 8 | 29±0.6 | |||
| Kanamycin | 30 | 16±0.3 | |||
| hBD2 | 1 | Salmonella spp.ISOLATE 6 | 17±0.5 | ||
| hBD2 | 4 | 22±0.5 | |||
| hBD2 | 8 | 26±0.4 | |||
| Kanamycin | 30 | 28±0.3 | |||
| hBD2 | 1 | Salmonella arizonaISOLATE 7 | 18±0.3 | ||
| hBD2 | 4 | 19±0.3 | |||
| hBD2 | 8 | 21±0.4 | |||
| Kanamycin | 30 | 33±0.4 | |||
 S.marcesens, (B) P. aeruginosa, and (C) A. baumannii, during treatment with serial dilutions of the recombinant peptide. The higher peptide concentration tested was at 8μg/mL. The test was performed three times on different days, and the means and standard deviations are indicated. A statistically significant difference was observed (p≥0.05). The minimum inhibitory concentrations (MIC) of defensin for each strain is considered as the lowest concentration required to inhibit growth>90% of the control. MIC values are indicated in square brackets, as determined after 24h of exposure to human beta defensin-2.")
Effect of human beta defensin-2 on bacterial growth. Optical density at different times of growth in Luria-Bertani medium was followed for three strains, (A) S.marcesens, (B) P. aeruginosa, and (C) A. baumannii, during treatment with serial dilutions of the recombinant peptide. The higher peptide concentration tested was at 8μg/mL. The test was performed three times on different days, and the means and standard deviations are indicated. A statistically significant difference was observed (p≥0.05). The minimum inhibitory concentrations (MIC) of defensin for each strain is considered as the lowest concentration required to inhibit growth>90% of the control. MIC values are indicated in square brackets, as determined after 24h of exposure to human beta defensin-2.
According to the CLSI, control of Kanamycin at a concentration of 30μg in the disk must produce an inhibition halo of 18-25mm against E.coli ATCC 25922.17 In the present study, the control showed an inhibition of 20mm (Table 1). There is no standard diameter measure for inhibition by defensin. However, strong bactericidal properties were observed against all seven isolates obtained from diarrheal stool with diameters over 18mm.
Additionally, hBD2 exhibited antimicrobial activity against three strains representative of bacterial species involved in nosocomial infections. MICs of 0.25μg/mL and 0.5μg/mL were observed for S. marcescen and P. aeruginosa, while a relatively higher MIC (4μg/mL) was observed against a clinical isolate of a multidrug-resistant strain of A. baumannii.
DiscussionDevelopmental delays in the immune system of human infants are compensated by the mammary gland, providing the necessary protection. Lactancy provides not only nourishment but also vital protection against environmental pathogens. Even with high interindividual variability in levels of different milk-related protecting factors, breastfeeding provides protection against necrotizing enterocolitis,4 gastrointestinal infection,3 and respiratory infections2 in the newborn. This article addressed the production of hBD2 in Venezuelan breast milk donors and explored the potential protective effect of hBD2 against enteropathogens isolates from children's diarrheal stool.
Wang et al. reported that hBD2 levels in Chinese Han mothers differed between colostrum and mature milk.18 In the colostrum, the concentration ranges of hBD2 were 0.31-19.12ng/mL, compared to 52.65-182.29pg/mL found in mature milk. In agreement with Wang et al., the present study also observed higher hBD2 levels in colostrum than in mature milk; however, both groups presented higher levels of hBD2 than the Chinese Han group.18 The differences between the hBD2 expression levels between both studies are striking, considering that both assessed an equal number of donors. Therefore, these differences could be attributed to the ethnicity of the groups involved. A recent article by Burch et al. highlighted that race/ethnicity appears to have an influence on the level of immune factors in breast milk;19 the present results appear to support these findings. To the best of the authors’ knowledge, this study is the first report of the characterization of defensin levels in human milk in Latin American women, and the present findings help to reinforce the recommendation of exclusive breastfeeding aiming to protect the health of at-risk children. Defensins have been found at a concentration of 1–10mg/mL in neutrophil granules20,21 and at concentration of 20–100mg/mL in porcine tongue epithelium.22 It is believed that lower concentrations (0.1–1mg/ml) of β-defensins might induce chemotaxis;23,24 the present results showcase a similar range of concentration.
In agreement with the present results, Castellote et al.25 reported that the concentration of immune factors in colostrum samples is richer than both transitional and mature milk. They suggested that the high content of active factors in premature infants and their inverse correlations to birth weight could indicate that their concentration in milk is modulated by the neonate's requirements. The mechanisms responsible for differences in the composition of milk between stages of lactation have yet to be established.
In vitro bactericidal activity of human-defensin-2 against enteropathogens and nosocomial strains has been reported.26 Indeed, each of the beta-defensins characterized to date has the ability to kill or inhibit in vitro a variety of bacteria. This activity varies particularly with concentrations of salt and plasma proteins in the assay.21 Thus, direct comparison of the MIC could be non-determinant. However, it is striking that the concentration at which this defensin shows antimicrobial activity is the same as human milk's, highlighting its physiological function.
The role of this peptide in the intestinal physiology of the newborn has been assumed as antimicrobial. Bacterial diarrhea is one of the main causes of infection in childhood. Therefore, it is important to determine the activity of hBD2 in isolates of diarrheic clinical cases. In the present study, four different isolates of E.coli and three of Salmonella spp. were used. In all cases, an inhibitory growth activity was observed when comparing with the reference parameters of the antibiotic kanamycin (with isolate referral strain E.coli ATCC 25922). Kapel et al.13 described elevated fecal levels of hBD2 in pediatric patients with both ulcerative colitis (UC) and Crohn's disease (CD) compared to healthy controls. They found a three-to-four fold increase in fecal hBD2 levels in children with CD as opposed to a>ten-fold increase in fecal hBD2 levels for UC. Notably, Barrera et al.11 reported that intestinal epithelial cells increased the secretion of hBD2 after treatment with catalytic antibodies with protease activity purified from breast milk. Therefore, it should be considered that the content of hBD2 in stool samples corresponds to the sum of the intestinal peptide production plus the direct intake through breast milk. Moreover, Hiratsuka et al.27 have indicated a reduced activity of beta-defensins in cystic fibrosis, suggesting that beta-defensins may play an important role in the pathological process of chronic respiratory tract infection. Besides its antimicrobial activity, other activities for hBD2 have been reported. Findings of Gambichler et al.28 showed altered expression patterns of hBD2 in non-ulcerated basal cell carcinoma, suggesting that hBD2 may also play a role in the pathogenesis of this non-melanoma skin cancer. Recently, Li et al.29 indicated that gene therapy with hBD2 could mediate specific antitumor immunity and increase local antitumor effects. Additionally, hBD2 has shown chemotactic activity for monocytes, macrophages, neutrophils, and immature dendric cells.15
Finally, the antimicrobial activity of recombinant hBD2 was determined over three Gram-negative bacteria (S. marcescen, P. aeruginosa, A. baumannii) that can be opportunistic pathogens in humans, affecting individuals with compromised immune systems. A major factor in their prominence as pathogens is their intrinsic resistance to antibiotics and disinfectants, which makes them important subjects in the field of nosocomial infection. Recombinant hBD2 showed inhibited growth in all tested bacteria. The hBD2 susceptibility showed by multidrug-resistant A. Baumannii strains is in agreement with that reported by Routsias et al.,26 who suggested that, besides mutations that confer resistance to certain antibiotics, this bacteria suffers mutations that affect energy generation required for maintenance of its membrane integrity or affect membrane structure itself, leading to hyper-susceptibility to hBD2. The lytic action of AMPs occurs through a binding event whereby the positively charged peptides interact with the anionic plasma membranes of targeted microorganisms. It has been known that the antimicrobial activity of hBD2 and other AMPs is highly sensitive to the presence of divalent cations, serum, and anionic macromolecules such as glycosaminoglycans. However, many of these AMPs are still able to confer protection under these conditions. In this regard, other immunomodulatory properties, such as cell migration, survival, proliferation, and induction of immune mediators, could probably be more significant in their disease-fighting mechanisms.30
The misuse of antibiotics has led to horizontal gene transfer among microbes and stimulated their evolutionary potential to develop resistance against conventional antimicrobials. In consequence, bacterial resistance to antibiotics is an escalating problem in modern medicinal treatment of infectious diseases, as shown by a growing number of immunocompromised patients with nosocomial infections. New therapeutic approaches are needed in order to temporarily overcome the resistance problem. Since antimicrobial peptides are products of long-term evolution in nature, the study of this peptide family could provide a feasible solution.
In summary, the female population in this study had hBD2 secreted in milk, suggesting that race/ethnicity could influence the level of immune factors in breast milk. High levels of peptide found in colostrum suggest the importance of this innate immunity factor during the first days of life. The hBD2 peptide showed antimicrobial activity against a range of bacteria, including common pathogens that cause diarrhea in infants and gram-negative relevant in nosocomial infections. The activity and high hBD2 content in breast milk highlight the superiority of breastfeeding in the nutrition of human infants.
Conflicts of interestThe authors declare no conflicts of interest.
This work was supported by a grant from Instituto Venezolano de Investigaciones Científicas (IVIC). This study was part of Joanna Baricelli's Master thesis at the Centro de Estudios Avanzados (CEA-IVIC).
Please cite this article as: Baricelli J, Rocafull MA, Vázquez D, Bastidas B, Báez-Ramirez E, Thomas LE. β-defensin-2 in breast milk displays a broad antimicrobial activity against pathogenic bacteria. J Pediatr (Rio J). 2015;91:36–43.
This work was supported by a grant from Instituto Venezolano de Investigaciones Científicas (IVIC), Caracas, Venezuela.




