
To analyze the association between the Trp64Arg polymorphism of the ADRB3 gene, maximal fat oxidation rates and the lipid profile levels in non-obese adolescents.
Methods72 schoolchildren, of both genders, aged between 11 and 17 years, participated in the study. The anthropometric and body composition variables, in addition to total cholesterol, HDL-c, LDL-c, triglycerides, insulin, and basal glycemia, were evaluated. The sample was divided into two groups according to the presence or absence of the polymorphism: non-carriers of the Arg64 allele, i.e., homozygous (Trp64Trp: n = 54), and carriers of the Arg64 allele (Trp64Arg + Arg64Arg: n = 18), in which the frequency of the Arg64 allele was 15.2%. The maximal oxygen uptake and peak of oxygen uptake during exercise were obtained through the symptom-limited, submaximal treadmill test. Maximal fat oxidation was determined according to the ventilatory ratio proposed in Lusk's table.
ResultsAdolescents carrying the less frequent allele (Trp64Arg and Arg64Arg) had higher LDL-c levels (p 0.031) and lower maximal fat oxidation rates (p = 0.038) when compared with non-carriers (Trp64Trp).
ConclusionsAlthough the physiological processes related to lipolysis and lipid metabolism are complex, the presence of the Arg 64 allele was associated with lower rates of FATMAX during aerobic exercise, as well as with higher levels of LDL‐c in adolescents.
Analisar a associação entre o polimorfismo Trp64Arg do gene ADRB3, as taxas de oxidação máxima de gorduras e as concentrações do perfil lipídico em adolescentes não obesos.
MétodosParticiparam do estudo 72 escolares, de ambos os sexos, com idade entre 11 e 17 anos. Foram avaliadas as variáveis antropométricas e de composição corporal, além do colesterol total, lipoproteina de alta densidade, lipoproteina de baixa densidade, triglicerídeos; insulina e glicemia basal. A amostra foi dividida em dois grupos, segundo a presença ou não do polimorfismo: não portadores do alelo Arg64, ou seja, homozigotos (Trp64Trp: n = 54) e portadores do alelo Arg64 (Trp64Arg + Arg64Arg: n = 18), em que a frequência do alelo Arg64 foide 15,2%. O consumo máximo de oxigênio e pico de consumo máximo de oxigênio durante o exercício foram obtidos por meio do teste aeróbio submáximo de sintoma limitado em esteira. A oxidação máxima de gorduras foi determinada de acordo com a razão de trocas ventilatórias propostas na Tabela de Lusk.
ResultadosOs adolescentes portadores do alelo menos frequente (Trp64Arg e Arg64Arg) apresentaram maiores concentrações de lipoproteina de baixa densidade (p = 0,031) e menores taxasde oxidação máxima de gorduras (p = 0,038) quando comparados aos não portadores (Trp64Trp).
ConclusõesEmbora os processos fisiológicos relacionados à lipólise e ao metabolismo de lipídeos sejam complexos, a presença do alelo Arg64 associou-se a menores taxas de FATMAX durante exercício aeróbio, bem como maiores níveis de lipoproteina de baixa densidade em adolescentes.
O exercício aeróbio é uma das principais estratégias na regulação do peso corporal, prevenção e tratamento da obesidade de origem complexa e diabetes mellitus tipo 2 (DM2), pois, além de potencializar o gasto energético, aumenta a absorção de glicose das células musculares e promove diminuição da resistência insulínica.1 No entanto, a população em geral não se beneficia igualmente da prática de exercícios físicos, visto que, além do próprio exercício aplicado, tais efeitos dependem de outros fatores ambientais, como a dieta e o background genético individual.2 É bem descrito o importante papel que genes específicos têm na regulação do peso por meio da ação de seus produtos no gasto energético, oxidação de substratos, modulação do apetite, metabolismo lipídico, termogênese e diferenciação celular.2,3
Nesse contexto, um dos sistemas mais importantes é o adrenérgico, que atua na regulação do equilíbrio de energia por meio da termogênese do tecido adiposo marrom e lipólise no tecido adiposo branco, tanto em humanos quanto em outras espécies.4,5 Parte desse sistema encontra‐se no receptor β3, localizado no cromossomo 8p 11.23, expresso principalmente no tecido adiposo visceral e subcutâneo, atua como mediador da lipólise e no tecido adiposo marrom como regulador da termogênese, ambas funções desempenhadas em resposta à estimulação por catecolaminas.6,7
Alterações da funcionalidade e da quantidade dos receptores β3 expressos podem permear diferenças individuais de gasto energético em resposta às atividades físicas. Entre algumas de suas variantes associadas às disfunções metabólicas, destaca‐se o polimorfismo de um único nucleotídeo (single nucleotide polymorphism – SNP) Trp64Arg (rs4994), que consiste na substituição de uma timina por uma citosina (T > C), que resulta na troca de um triptofano por uma arginina na posição 64da proteína madura e corresponde ao primeiro loop intracelular do receptor β3.8 Portadores do alelo Arg64 se mostraram mais resistentes à perda de peso e à diminuição de gordura visceral,9 além de ter maiores chances de apresentar alterações lipídicas, obesidade, DM27 e taxas reduzidas de oxidação de gordura.10
Tanto os processos metabólicos normais que culminam na oxidação de gordura quanto as alterações metabólicas relacionadas ao perfil lipídico são sistemas complexos que envolvem múltiplas vias e interações de fatores exógenos (ambientais) e endógenos.11
Apesar da complexidade subjacente a esses processos, estudos têm procurado identificar a pequena parcela que pode ser atribuída a variantes alélicas de genes cujos produtos podem influenciar nos processos fisiológicos supracitados. Dessa forma, o presente estudo tem por objetivo verificar a associação entre o polimorfismo Trp64Arg, as taxas de oxidação máxima de gorduras e as concentrações do perfil lipídico em adolescentes não obesos.
MétodosDelineamento experimentalEstudo transversal e observacional, caracterizado por triagem antropométrica inicial de todos os adolescentes de escola pública e urbana do Sul do Brasil. Participaram da pesquisa 1.077 escolares, de ambos os sexos e entre 11 e 17 anos. A prevalência de adolescentes não obesos classificados entre o percentis 5° e 97°, conforme proposto pela Organização Mundial de Saúde,12 foi de 93%. Na sequência, todos os alunos foram convidados a participar de um programa de atividades físicas, em que 165 sujeitos foram voluntários. Os participantes foram avaliados por uma única médica pediátrica com anamnese completa, exames antropométricos e análise da maturação sexual conforme proposto por Tanner.13
Os critérios de inclusão foram: ausência de doenças crônicas como DM2, hipotireoidismo não controlado, doenças endócrinas, infecções e/ou uso de medicamentos que promovam alterações na adiposidade, parâmetros metabólicos e inflamatórios. O critério de exclusão adotado foi a classificação do estado nutricional obeso (n = 85), considerados o percentil acima de 97° pela Organização Mundial da Saúde.12 Todos os voluntários e os pais assinaram o termo de consentimento livre e esclarecido, conforme projeto de pesquisa aprovado por Comitê de Ética Institucional (protocolo n°. 2460.067/2011‐03‐UFPR).
Após aplicação dos critérios de inclusão e exclusão, a seleção resultou em 80 adolescentes não obesos (38 meninos e 42 meninas), os quais não faziam mais de 120 minutos/semana de atividade física e que não apresentaram contraindicação para os exames, inclusive ausência de doenças cardíacas, pulmonares e osteoarticulares, foram submetidos ao teste de esforço submáximo em esteira e coletas de sangue. Desses, cinco adolescentes não fizeram os testes de avaliação cardiorrespiratória e em três não foi possível fazer a genotipagem do polimorfismo Trp64Arg do gene ADRB3, resultou em 72 adolescentes não obesos (33 meninos e 39 meninas).
Avaliação das variáveis antropométricas e fisiológicasMassa corporal (MC) (kg) foi obtida em balança tipo plataforma, com precisão de 0,1 quilograma (kg) e capacidade máxima de 150kg. A estatura foi aferida em estadiômetro fixado à parede, com precisão de 0,1cm e amplitude de 220cm. Medida da circunferência abdominal (CA) foi avaliada criteriosamente de acordo com as recomendações do Centers for Disease Control and Prevention.14
Avaliação da composição de gordura corporal foi feita com a absortiometria de raios X de dupla energia (DXA), com aparelho da marca Lunar Prodigy Primo (General Electric, Healthcare, MA, EUA). Massa magra (MM) e massa gorda (MG) foram determinadas com aproximação de 1g e descritas em quilogramas (kg). Para reconstrução da imagem da MM e MG foi usado o software enCore 2008 versão 12.30 (General Electric, Healthcare, MA, EUA).
O consumo máximo de oxigênio (VO2max) e o pico do consumo máximo de oxigênio (VO2pico) foram mensurados em esteira ergométrica por meio do analisador de gases portátil K4b2® (Cosmed®, SP, Brasil), com uso do seguinte protocolo: velocidade inicial de 4km/h, aumento progressivo de 0,3km/h a cada 30 segundos e inclinação fixa de 1% mantida constante durante o teste. Frequência cardíaca (FC) foi monitorada durante o teste, por meio de cardiofrequencímetro (Polar®, EUA), sistema portátil de recepção‐transmissão wireless. Os adolescentes foram orientados por nutricionista para fazer alimentação leve com predominância de carboidratos antes do teste, segundo Recomendações Diárias Alimentares.15 O teste foi considerado máximo quando dois dos seguintes critérios foram observados: a) exaustão ou inabilidade para manter velocidade requerida; b) R ≥ 1,09; c) alcançar a FC máxima prevista pela fórmula 208 – (0,7 x idade), proposta por Tanaka.16
Para obtenção dos valores da oxidação máxima de gorduras (Fatmax) foram usadas razões de troca ventilatória (R) observadas durante teste aeróbio submáximo de sintoma limitado, conforme tabela de Lusk.17 Essa variável foi obtida por meio do produto entre o equivalente calórico do R no ponto da Fatmax e consumo de oxigênio observado nesse mesmo ponto.
Avaliação das variáveis bioquímicasAmostras sanguíneas foram coletadas no período da manhã após 12 horas de jejum e acondicionadas em tubos apropriados. Concentrações plasmáticas de colesterol total, HDL colesterol e triglicerídeos foram determinadas em mg/dl, através de teste colorimétrico enzimático. LDL colesterol foi calculado pela equação de Friedewald em mg/dl.18 Valores de glicemia foram determinados pelo método enzimático (Glucose Oxidase – Labtest, SP, Brasil) e insulina dosada pela técnica de quimiluminescência por imunoensaio imunométrico em uU/ml, em equipamento automatizado.
Genotipagem do polimorfismo Trp64Arg do gene ADRB3A extração de DNA das amostras de sangue foi feita pelo QIAamp DNA Mini Kit (QIAGEN, MD, EUA), conforme método de Lahiri & Nurnberger.19 A genotipagem do polimorfismo Trp64Arg do gene ADRB3 foi feita por meio do ensaio de discriminação alélica TaqMan com o equipamento 7500TMreal time PCR system (Applied Biosystems®, CA, EUA), as reações foram avaliadas em um aparelho Eppendorf Matercycler Realplex 2 (Biocompare®, CA, EUA) e as análises feitas com o software Eppendorf realplex v. 1.5 (Biocompare®, CA, EUA).
A extração de DNA constituiu‐se na obtenção da camada de leucócitos por centrifugação do sangue total coletado, lise de eritrócitos, precipitação de proteínas e precipitação de DNA genômico. A técnica de PCR permitiu a detecção, ciclo a ciclo, com elevada sensibilidade e especificidade, da intensidade de fluorescência emitida em decorrência da amplificação da sequência de DNA‐alvo. No fim da reação, de acordo com o genótipo, apresentou‐se no caso dos homozigotos uma ou outra fluorescência e no caso de heterozigotos ambas as fluorescências. A distribuição dos genótipos entre os adolescentes que compuseram o estudo foi: Trp/Trp (n = 54) (75%), Trp/Arg (n = 14) (19,5%) e Arg/Arg (n = 4) (5,5%). A frequência do alelo do Arg64 foi de 15,2%, indicou que os dados estão em equilíbrio de Hardy‐Weinberg, ou seja, os valores observados são semelhantes aos esperados (p = 0,165). Os modelos de interação alélica recessivo, dominante e com ausência de dominância foram testados por adotar o modelo dominante de interação alélica, devido ao número reduzido de portadores do alelo Arg64 em homozigose e também devido ao efeito observado. Assim, os indivíduos foram agrupados em portadores (Arg/Arg + Trp/Arg) e não portadores (Trp/Trp) do alelo Arg64.
Análises estatísticasOs dados foram testados quanto à normalidade e homogeneidade de variâncias pelos testes de Kolmogorov‐Smirnov e Correlação de Significância de Lilliefors, respectivamente. Os resultados são apresentados em médias e desvio‐padrão para as variáveis com distribuição normal e medianas e intervalos interquartis para as não normais. Usou‐se teste t de Student para variáveis com distribuição normal e o teste U de Mann‐Whitney para variáveis que não atenderam ao pressuposto de normalidade. O teste exato de Fisher foi adotado para comparações de proporções entre grupos quanto ao estágio maturacional. Para análise da correlação entre as variáveis bioquímicas e a máxima oxidação de gorduras foi usada a correlação de Spearman. As frequências genotípicas foram obtidas por contagem direta, as comparações de proporções entre os grupos quanto ao sexo e o equilíbrio de Hardy‐Weinberg foram verificadas por meio do teste de qui‐quadrado (x2). A regressão linear múltipla foi usada para verificar o grau de associação entre maturação sexual, sexo, massa gorda e polimorfismo Trp64Arg do gene ADRB3 (variáveis independentes), perfil lipídico e oxidação máxima de gorduras (desfecho). Calculou‐se a posteriori o poder do estudo no software G*power3 (G Power, V.3.1.7, Dusseldorf, Alemanha) com a presente amostra e identificou‐se um poder (1‐β) de 0,66 para as comparações entre grupos, de 0,84 para as correlações e de 0,82 para as regressões. Os dados foram analisados com programa SPSS for Windows (IBM SPSS Statistics para Windows, versão 22.0. NY, EUA) com nível de significância de p < 0,05.
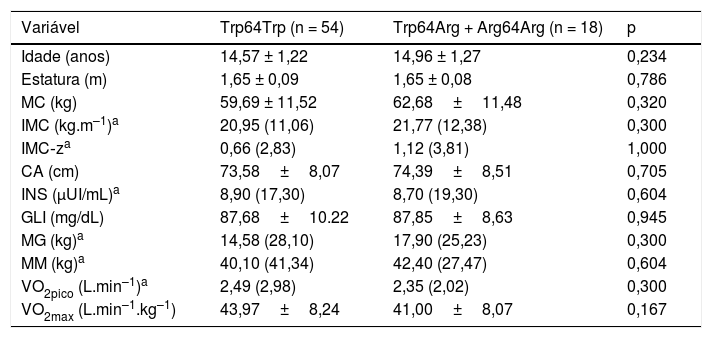
ResultadosA tabela 1 apresenta as variáveis antropométricas, de composição corporal, bioquímicas e aptidão cardiorrespiratória dos adolescentes não portadores (Trp64Trp) e portadores do alelo menos frequente (Trp64Arg + Arg64Arg), que foram semelhantes. Não houve diferença estatística significativa entre os grupos quanto aos estágios de Tanner.
Comparação das médias ± desvio‐padrão ou medianas (intervalos de confiança) das variáveis antropométricas, de composição corporal, bioquímicas e de aptidão cardiorrespiratória entre adolescentes não portadores (Trp64Trp) e portadores do alelo menos frequente (Trp64Arg + Arg64Arg) do gene ADRB3
| Variável | Trp64Trp (n = 54) | Trp64Arg + Arg64Arg (n = 18) | p |
|---|---|---|---|
| Idade (anos) | 14,57 ± 1,22 | 14,96 ± 1,27 | 0,234 |
| Estatura (m) | 1,65 ± 0,09 | 1,65 ± 0,08 | 0,786 |
| MC (kg) | 59,69 ± 11,52 | 62,68±11,48 | 0,320 |
| IMC (kg.m–1)a | 20,95 (11,06) | 21,77 (12,38) | 0,300 |
| IMC‐za | 0,66 (2,83) | 1,12 (3,81) | 1,000 |
| CA (cm) | 73,58±8,07 | 74,39±8,51 | 0,705 |
| INS (μUI/mL)a | 8,90 (17,30) | 8,70 (19,30) | 0,604 |
| GLI (mg/dL) | 87,68±10.22 | 87,85±8,63 | 0,945 |
| MG (kg)a | 14,58 (28,10) | 17,90 (25,23) | 0,300 |
| MM (kg)a | 40,10 (41,34) | 42,40 (27,47) | 0,604 |
| VO2pico (L.min–1)a | 2,49 (2,98) | 2,35 (2,02) | 0,300 |
| VO2max (L.min–1.kg–1) | 43,97±8,24 | 41,00±8,07 | 0,167 |
CA, circunferência abdominal; GLIC, glicemia basal; IMC, índice de massa corporal; IMC‐z, índice de massa corporal escore Z; INS, insulina basal; MC, massa corporal; MG, massa gorda; MM, massa magra.
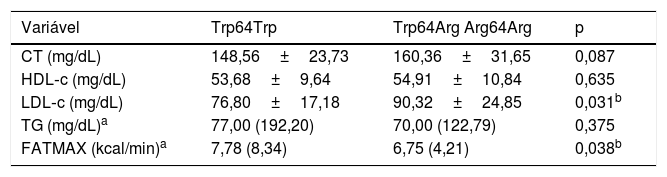
A tabela 2 demonstra que o grupo portador do alelo Arg64 apresentou maiores concentrações de LDL‐C em comparação com o grupo não portador (p = 0,031). As concentrações de CT, HDL‐c e TG foram semelhantes entre os grupos. O grupo portador do alelo Arg64 apresentou menores taxas de Fatmax (p = 0,038) quando comparado com o grupo não portador (Trp64Trp).
Comparação das médias±desvio‐padrão ou medianas (intervalos de confiança) das variáveis do perfil lipídico e da oxidação máxima de gorduras entre adolescentes não portadores (Trp64Trp) e portadores do alelo menos frequente (Trp64Arg + Arg64Arg) do gene ADRB3
| Variável | Trp64Trp | Trp64Arg Arg64Arg | p |
|---|---|---|---|
| CT (mg/dL) | 148,56±23,73 | 160,36±31,65 | 0,087 |
| HDL‐c (mg/dL) | 53,68±9,64 | 54,91±10,84 | 0,635 |
| LDL‐c (mg/dL) | 76,80±17,18 | 90,32±24,85 | 0,031b |
| TG (mg/dL)a | 77,00 (192,20) | 70,00 (122,79) | 0,375 |
| FATMAX (kcal/min)a | 7,78 (8,34) | 6,75 (4,21) | 0,038b |
CT, colesterol total; Fatmax, oxidação máxima de gorduras; HDL‐c, lipoproteína de alta densidade; LDL‐c, lipoproteína de baixa densidade; TG, triglicerídeos.
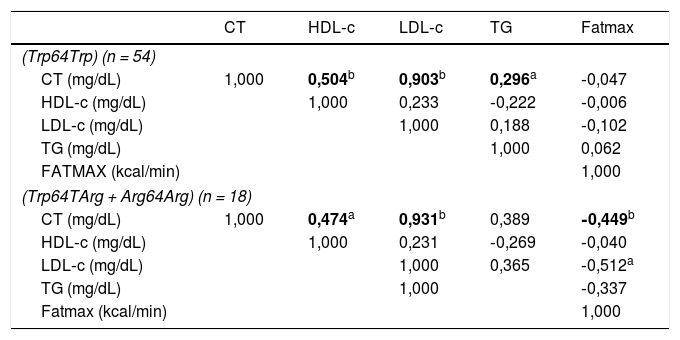
A tabela 3 apresenta valores de correlação de Spearman em que os níveis de HDL‐c (p = 0,000; r = 0,504), LDL‐c (p = 0,000; r = 0,903) e TG (p = 0,035; 0,296) dos sujeitos não portadores do alelo Arg64 foram diretamente correlacionados com o colesterol total (CT). Entre os portadores do alelo Arg64 a Fatmax mostrou correlação indireta com as concentrações de LDL‐c (p = 0,018; r = 0,512) e CT (p = 0,041; r = 0,449). Além disso, os níveis séricos de HDL‐C (p = 0,030; r = 0,474) e LDL‐C (p = 0,000; r = 0,931) correlacionaram‐se diretamente com o colesterol total. Não houve correlações significativas entre demais variáveis.
Matriz dos valores dos coeficientes de correlação de Spearman entre as variáveis do perfil lipídico e a oxidação máxima de gorduras nos adolescentes homozigotos para o alelo mais frequente (Trp64Trp) (n = 54) e para os portadores do alelo menos frequente (Trp64TArg + Arg64Arg) (n = 18)
| CT | HDL‐c | LDL‐c | TG | Fatmax | |
|---|---|---|---|---|---|
| (Trp64Trp) (n = 54) | |||||
| CT (mg/dL) | 1,000 | 0,504b | 0,903b | 0,296a | ‐0,047 |
| HDL‐c (mg/dL) | 1,000 | 0,233 | ‐0,222 | ‐0,006 | |
| LDL‐c (mg/dL) | 1,000 | 0,188 | ‐0,102 | ||
| TG (mg/dL) | 1,000 | 0,062 | |||
| FATMAX (kcal/min) | 1,000 | ||||
| (Trp64TArg + Arg64Arg) (n = 18) | |||||
| CT (mg/dL) | 1,000 | 0,474a | 0,931b | 0,389 | ‐0,449b |
| HDL‐c (mg/dL) | 1,000 | 0,231 | ‐0,269 | ‐0,040 | |
| LDL‐c (mg/dL) | 1,000 | 0,365 | ‐0,512a | ||
| TG (mg/dL) | 1,000 | ‐0,337 | |||
| Fatmax (kcal/min) | 1,000 | ||||
CT, colesterol total; Fatmax, oxidação máxima de gorduras; HDL‐c, lipoproteína de alta densidade; LDL‐c, lipoproteína de baixa densidade; TG, triglicerídeos.
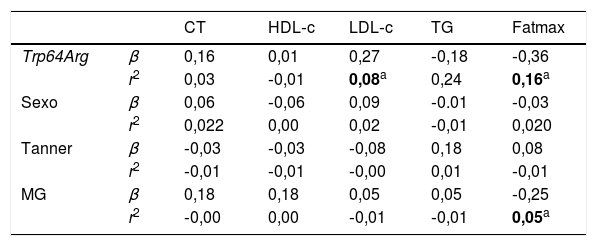
O teste de regressão linear múltipla (tabela 4) mostrou que o polimorfismo Trp64Arg do gene ADRB3 explicou 8% (r2 ajustado = 0,08) dos níveis de LDL‐c (β = 0,27, p < 0,01) e 16% (r2 ajustado = 0,16) da Fatmax (β = ‐0,36, p < 0,01), enquanto que a MG explicou 5% (r2 ajustado = 0,05) da Fatmax (β = ‐0,25, p < 0,01).
Coeficiente de regressão linear (β) e r2 para as variáveis de sexo, maturação sexual, polimorfismo Trp64Arg do gene ADRB3, perfil lipídico e oxidação máxima de gorduras em adolescentes
| CT | HDL‐c | LDL‐c | TG | Fatmax | ||
|---|---|---|---|---|---|---|
| Trp64Arg | β | 0,16 | 0,01 | 0,27 | ‐0,18 | ‐0,36 |
| r2 | 0,03 | ‐0,01 | 0,08a | 0,24 | 0,16a | |
| Sexo | β | 0,06 | ‐0,06 | 0,09 | ‐0.01 | ‐0,03 |
| r2 | 0,022 | 0,00 | 0,02 | ‐0,01 | 0,020 | |
| Tanner | β | ‐0,03 | ‐0,03 | ‐0,08 | 0,18 | 0,08 |
| r2 | ‐0,01 | ‐0,01 | ‐0,00 | 0,01 | ‐0,01 | |
| MG | β | 0,18 | 0,18 | 0,05 | 0,05 | ‐0,25 |
| r2 | ‐0,00 | 0,00 | ‐0,01 | ‐0,01 | 0,05a |
CT, colesterol total; Fatmax, oxidação máxima de gorduras; HDL‐c, lipoproteína de alta densidade; LDL‐c, lipoproteína de baixa densidade; TG, triglicerídeos.
No presente trabalho o objetivo foi investigar a associação do polimorfismo Trp64Arg sobre a oxidação máxima de gorduras e as variáveis bioquímicas em adolescentes não obesos. Tais parâmetros fisiológicos apresentam grande variação na população em geral, inclusive entre os adolescentes, em função da interação entre múltiplos fatores genéticos determinantes, e desses com componentes ambientais, tais como hábitos alimentares e estilo de vida.
Os resultados sugerem que as taxas de Fatmax foram significativamente menores e os níveis de LDL‐c foram maiores entre os adolescentes portadores do alelo Arg64, quando comparados com os adolescentes com a ausência do alelo. Resultados semelhantes são observados em adultos eutróficos, tanto em repouso quanto durante o exercício aeróbio, em que o polimorfismo Trp64Arg foi associado à redução da oxidação máxima de gorduras.10 Ainda, nesse sentido, pesquisa feita in vitro com adipócitos humanos sugeriu atividade lipolítica reduzida em indivíduos homozigotos portadores do alelo Arg64.20 Embora seja um processo fisiológico complexo, a variante genética do gene ADRB3 pareceu contribuir na variação dos valores de Fatmax em resposta à atividade física, bem como pode contribuir para uma menor capacidade de resposta à perda de peso.8
A prescrição de atividades físicas no ponto da oxidação máxima de gorduras é um importante fator que pode contribuir para a melhoria do metabolismo lipídico e a manutenção do peso,21 bem como o aumento de sensibilidade insulínica.22 O treinamento na zona da Fatmax sugere aumentos significativos na oxidação máxima de gorduras, conforme observado em adolescentes obesas que também apresentaram alterações favoráveis nas concentrações de adipocitocinas.23 Assim, compreender os fatores que contribuem para a variação individual das taxas de Fatmax pode possibilitar a prescrição de treinamentos mais individualizados que refletirão em resultados mais positivos.
Embora o presente estudo não tenha contemplado ensaios funcionais a fim de demonstrar o efeito da alteração no funcionamento das vias desencadeadas pela ativação do receptor β3, estudos anteriores demonstraram sua funcionalidade. Pesquisa feita em ratos comprovou que o portador do alelo Arg64 apresentou redução da quantidade de cAMP em resposta a catecolaminas, quando comparada com a presença do alelo 64Trp.24 O mesmo efeito também foi observado em outras células (células de ovário em hamsters e células renais embrionárias humanas),4,5 sugeriu que a lipólise pode estar diminuída na presença da variante Arg64 nos receptores β3 comparado com o receptor sem a alteração. Ainda nesse sentido, tal alteração genética pode refletir em um perfil metabólico menos favorável ao associar‐se com a termogênese do tecido adiposo marrom, ao peso corporal e ao início precoce da DM2 em algumas populações.6,7
Este estudo aponta que os portadores do alelo Arg64 apresentaram maiores concentrações de LDL‐c, o que pode representar maiores riscos para o desenvolvimento de doenças cardiovasculares no futuro. Da mesma forma, pesquisa feita com japoneses adultos e eutróficos apontou correlação significativa entre os níveis de LDL‐c, o genótipo Arg64Arg e a idade, com o aumento anual do IMC.25 Entretanto, na literatura é possível observar heterogeneidade de resultados. Em adultos italianos e não obesos não foi encontrada influência do alelo Arg64 sobre os níveis lipídicos. No entanto, tal alelo parece influenciar na adiposidade abdominal.26 Estudo em crianças húngaras de variados estados nutricionais não encontrou influência da presença do alelo Arg64 nas concentrações de TG, CT, HDL‐c e glicemia, porém os autores observaram maiores valores de massa corporal, massa gorda e insulina em jejum, quando comparados com os não portadores do alelo.27 Em outro estudo, indivíduos homozigotos Arg64Arg apresentaram secreção de insulina significativamente menor e níveis de glicose em jejum superiores, quando comparados com homozigotos Trp64Trp com excesso de peso.7
Ainda, os resultados sugerem que as maiores taxas de oxidação máxima de gorduras no grupo não portador do alelo Arg64 do gene ADRB3 apresentaram correlação negativa com as concentrações de CT e HDL‐c. Tal relação é conhecida, uma vez que o aumento do gasto energético leva a alterações no perfil lipídico secundário ao exercício físico, que acontecem pelo estímulo das reações metabólicas e pela potencialização do uso de substratos energéticos pela musculatura ativa, que ocorre tanto de forma aguda quanto por adaptações fisiológicas que estimulam o metabolismo ao longo do dia.21 Além disso, embora tenha‐se observado nas regressões o polimorfismo Trp64Arg do gene ADRB3 associado aos níveis de LDL‐c e Fatmax, deve ser levado em consideração o efeito de variantes genéticas nos traços complexos em tais processos fisiológicos, que podem ser atribuídos à faixa etária e ao estado nutricional dos sujeitos, bem como à heterogeneidade étnica do background genético.
O alelo Arg64 é encontrado em diferentes frequências entre as etnias estudadas e no presente estudo a frequência encontrada foi de 15,3%. Pesquisas em outras populações relataram 5,77% das crianças obesas húngaras,7 8% em euro‐americanos25 e em 11% em adultos brasileiros com diferentes estados nutricionais.28
O presente estudo apresenta algumas limitações que devem ser mencionadas. Primeiro, caracteriza‐se como delineamento transversal, por isso não foi possível estabelecer relação de causa e efeito entre a presença do polimorfismo, a oxidação máxima de gorduras e o perfil lipídico, se considerarmos que essa condição é multifatorial. Segundo, o número amostral reduzido (n = 72) impossibilitou a análise individualizada pelo sexo. Dessa forma, a generalização dos resultados obtidos com este trabalho deve ser usada com cautela, sabe‐se da influência que o sexo pode ter nesse contexto. Terceiro, o uso de uma amostra de conveniência torna a homogeneidade entre os grupos selecionados difícil de ser alcançada, pois diferenças no sexo e estágio de Tanner poderão alterar as taxas de Fatmax e os níveis do perfil lipídico. Entretanto, nenhuma diferença foi encontrada nas comparações entre os sexos e o estágio de Tanner nos grupos divididos pela presença ou ausência do alelo Arg64. Contudo, outros aspectos fortalecem nossos achados, como o método usado para a análise da composição da gordura corporal (DXA) – considerado gold‐standard –, os critérios de inclusão e exclusão rigorosos e o uso protocolo de Lusk na avaliação da oxidação máxima de gorduras, o qual apresenta boa concordância com técnicas invasivas, como canulação arterial e biópsias musculares, além do fácil acesso e aplicabilidade pelos profissionais da área da saúde.29
A identificação da influência dos polimorfismos em genes candidatos sobre as taxas de oxidação máxima de gorduras e metabolismo lipídico pode contribuir para o estabelecimento de protocolos de exercícios físicos mais individualizados e eficazes, capazes de contribuir de forma mais efetiva para o provimento da saúde. No entanto, a associação entre o alelo Arg64 do polimorfismo Trp64Arg do gene ADRB3, a oxidação máxima de gorduras e as concentrações de colesterol LDL deve ser interpretada com cautela, pois pode representar uma pequena parcela em processos fisiológicos complexos relacionados à lipólise e ao metabolismo de lipídeos.
FinanciamentoCoordenação de Aperfeiçoamento de Pessoal de Nível Superior (Capes) e Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Conflitos de interesseOs autores declaram não haver conflitos de interesse.
Como citar este artigo: Jesus ÍC, Alle LF, Munhoz EC, Silva LR, Lopes WA, Tureck LV, et al. Trp64Arg polymorphism of the ADRB3 gene associated with maximal fat oxidation and LDL‐C levels in non‐obese adolescents. J Pediatr (Rio J). 2018;84:425–31.
Trabalho vinculado à Universidade Federal do Paraná (UFPR), Curitiba, PR, Brasil.




